Journal of Unification Studies Vol. 12, 2011 - Pages 143-162
|
What missions must the new truth fulfill? The new truth should be able to unify knowledge by reconciling the internal truth pursued by religion and the external truth pursued by science. Consequently, it will enable all people to overcome the two types of ignorance, internal and external, and fully comprehend the two types of knowledge. –Exposition of the Divine Principle, Introduction |
As far as evolutionary biology is concerned, there are two main differences between Materialism and Unification Thought.
- In materialism, the organisms, species, genera, kingdoms, etc. that emerge over geological time are fortuitous and do not have a purpose or direction. This is expressed in the often-used aphorism: "Evolution has no foresight." In Unification Thought, the body plans and sequence of evolutionary emergence are designed and planned by God and follow a logical sequence of development leading up to the emergence of human beings.
- In materialism, evolution occurs by random variation in the genetic heritage of a lineage followed by natural selection of the variations most suited to survival and reproductive success. In Unification Thought, evolution occurs by the stepwise expression of the Logos over time.
In scientific terms, just exactly how the Logos has driven evolution along the path to Man has not been made clear in either the Divine Principle or in Unification Thought. This paper will attempt to start the dialog necessary to remedy this deficiency.
While this paper will be written mainly in the language of science, we will start with a few of the fundamental philosophical principles of Unifica¬tion Thought that will be harmonized with the scientific description.
Unification Principles
- From the inception of Creation in the Big Bang to the emergence of human beings, the physical universe has been in the Indirect Dominion of God. In this period of thirteen billion years or so, the universe has been ruled by the Logos, which was created by God before the creation was initiated. God has no direct control or input into the course of events during this time. The Direct Dominion of God only begins with the emergence of humans with the capacity for True Love.
- The physical universe was created and designed to be under the direct dominion of human beings so that they could create themselves and stand, as co-creators, as the children of God; not as robots or servants. This principle of "they did it themselves" has heretofore been considered to apply only to humans. In this work, we shall present the concept that this principle permeates the evolution of all life, and is not something new added at the inception of humans.
- The culmination of the Logos is the emergence of the form and capacity of the pre-human species that eventually gave birth to Adam and Eve. As the exact nature of this pre-human species is still an open question, we will refer to the biological parents of Adam and Eve as the Neanderthals as they are the best known to science so far. Going beyond the extent of the Logos, Adam and Eve should have grown through a period of responsibility to become True Humans with the same capacity for freely giving love as God.
- The inheritance of the merit of the ancestors—and its converse as the inheritance of sin—is a key principle in the Divine Principle and Unifi¬cation Thought. While this principle of inherited wisdom is usually only thought to apply to the human spirit, in this work we will present the concept that this Principle has been active since the origin of life and is the key to understanding the incremental process by which the Logos has been progressively expressed in the evolutionary process as designed by God.
It is not the intent of this work, however, to impose these philosophical principles on science; rather we shall show how they are being progressively expressed in the emerging science of epigenetics along with reasonable speculations as to how this discipline will develop in the future.
Logos and Origins
The Four Position Foundation principle as explained in Unification Thought is quite clear about how novel qualities of form and function become established in the inorganic realm. In these simple systems, the Origin of the first of a kind is essentially the same as the origin of all those that come after that first event. For example, the Origin of the first hydrogen atom in the universe (and there was a First) is similar to the origin of all the hydrogen atoms that make up 75% of the regular matter in the physical universe (helium atoms are 24% while all the other elements are just 1%).
For the first 300,000 years after the Big Bang the universe as a whole was much too hot for atoms to form—the overwhelming preponderance of gamma, X ray and ultraviolet photons of light over matter (100 billion photons to each particle of matter) ensured that electrons and protons could never get together in a relationship—the early universe was a hot plasma of free electrons, protons and helium nuclei. As the expansion of the universe continued and these high-energy photons became 'stretched' into low-energy photons of visible light, infrared and ultimately the Cosmic Microwave Background radiation of today.
At some point in time, an electron and proton combined together into the first atom of hydrogen—the Origin even—and then trillions of others soon emerged in the same way. Much, much later, after the first generation of massive stars had nucleo-burned their hydrogen fuel into oxygen and exploded as supernova, the first water molecule emerged when hydrogen and oxygen got together rapidly followed by many more.
To a materialist, it is happy chance that a hydrogen atom has the useful properties that it has, yet another even more fortuitous happy chance that the water molecule has the unique properties so necessary for life. The technical term for the properties of the hydrogen atom not present in isolated electrons and protons, or the properties of the water molecule not present in isolated hydrogen atoms or oxygen atoms is 'emergent properties,' a useful catchall term for all such fortuitous characteristics.
Unification Thought is more realistic; the fabulous properties that emerge when simple things get into complex relationships are the result of the careful design put into the Logos by the creator, our Heavenly Father.
The Logos does not direct the path of history of electrons and protons (or hydrogen atoms and oxygen atoms) so that they will hook up with each other. When they do encounter each other, however, the relationship they establish resonates with an aspect of the Logos (it is governed by natural law a scientist would say) and an intangible quality in the Logos becomes tangible in a physical system.
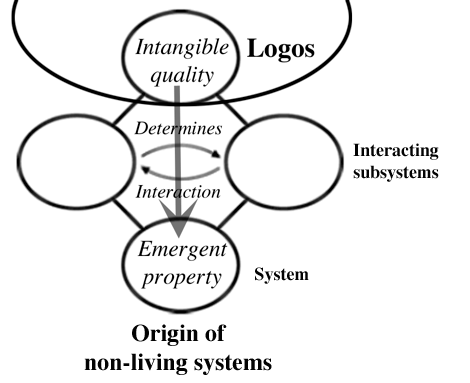
As this process involves waves, shapes and forms, we will call it 'analog learning from the Logos' as this will prevent a proliferation of terminology later on even though the sequence does not involve any sort of memory. For in inorganic systems, the Origin of the first specimen of a system is similar to the origin of all the similar systems that appear later on in creation.
For living systems, however, the Origin of the first of a type of system is fundamentally different from the origin of all those that emerge later. This is because there is an extra step involved; that of digital memory. Put simply, in living systems a novel analog form has its Origin in a relationship that resonates with the Logos (similar to the step in inorganic systems); a record of this new form is committed to digital memory; copies of the living system have their origin in the recall of this digital memory. It is this digital storage of Logos-derived information that differentiates living from non-living systems.
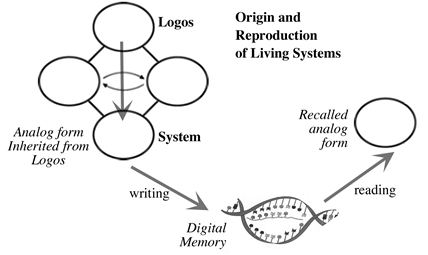
In this view, the random mistakes or 'mutations' that are so central to Darwinism play no constructive role in evolution. The Logos also has a role in the lives of the descendants: when they have a structure that resonates with the Logos we have health and prosperity; when the structure does not, we have illness and disease. This viewpoint places the Logos in a central role, unlike the simple 'fitness to survive and reproduce' that characterizes Darwinism.
The difference between the Origin of the first humans and the origin of all their descendants is fully-supported by the Divine Principle. While Adam and Eve were born sinless as a perfect reflection of the Logos, all their descendants (with just two exceptions) have been born imprinted with Original Sin, which is certainly not a reflection of any quality inherent in the Logos. This quality that does not reflect the Logos was written into genetic memory and passed on to all human beings.
|
Does original sin result from eating a fruit? It is said that the parents' eating of the fruit was the sin, but what is that fruit through which the descendants of thousands of generations have become sinners? This has to do with the blood relationship. If the root of sin is planted through lineage, it lasts forever by virtue of the law of heredity.[1] |
Epigenetics
While epigenetics is now so well established as to have a recent Nova episode[2] devoted to it on PBS television, it is probably a field that is unfamiliar to most people. In many ways, it can be considered as the reemergence of Lamarckism in a much more sophisticated form.
|
For years, genes have been considered the one and only way biological traits could be passed down through generations of organisms. Not anymore. Increasingly, biologists are finding that non-genetic varia¬tion acquired during the life of an organism can sometimes be passed on to offspring—a phenomenon known as epigenetic inheritance. An article... in the July issue of The Quarterly Review of Biology lists over 100 well-documented cases of epigenetic inheritance between generations of organisms, and suggests that non-DNA inheritance happens much more often than scientists previously thought.[3] |
The "central dogma" of molecular biology is that there is a one-way flow of information from the genotype—the genes and DNA sequence—to the phenotype—the proteins and the result of protein action, the development and eventual form and function of the body. It is upon this central dogma that the whole of Darwinism is constructed since, as there is no back-flow of information from the body to the genome, the only changes allowed in the genome are random mutations, random rearrangements and other such random occurrences for natural selection to go to work on.
This dogmatic assertion so fundamental to Darwinism is clearly up for revision. Note that the preeminent proponent of materialistic Darwinism, Richard Dawkins, assumes in his many works that all is now understood of the basic principles of evolution based on random mutation and variation.[4] As he does not, however, mention epigenetics even once in any of his writings, by this fact alone he his condemned to have only a partial view of the truth; the classic mistake of the blind man confusing his odiferous grasp of the elephant's tail with the whole beast. Richard Dawkins is not unique in this respect; it happened to many elder statesmen in physics just a century ago:
|
It seems that every so often, a fairly large group of scientists begin to assert that science is just about complete, that the vast unknown is gone, and that all the really major research can stop because we now know everything except the details. For those who fall under the spell of this sort of belief, be aware that a similar belief seemed to have taken hold at the turn of the last century. This was just before Relativity and Quantum Mechanics appeared on the scene and opened up new realms for exploration.... |
Just as in physics, where the advent of relativity and quantum mechanics punctured this "we know it all" attitude, so the advent of epigenetics has the potential to puncture the biological "we know it all" attitude prevalent in current Darwinism as exemplified by Richard Dawkins in all his writings. One can only feel sorry for Dawkins as the dustbin of history is not a comfortable place for one so arrogant.
Emergence of Epigenetics
The first hint that that the one-way 'central dogma' of Darwinism was wrong came when it was noticed that the identical genetic defect in the human genotype had very different effects on the phenotype depending on whether the disease was inherited from the mother or the father.
Even though both parents contribute equally to the genetic content of their offspring, a developmental process called genomic imprinting sometimes leads to the exclusive expression of specific genes from only one parent. This process was first described in 1984, when two laboratories discovered a mark, or "imprint," that differentiates between certain genes on the maternal and paternal chromosomes and results in the expression of only one copy of those genes in the offspring. The genes in imprinted areas of an organism's genome are expressed depending on the parent of origin.[6]
This phenomenon was eventually traced to a pattern of chemical alterations—methylation of the cytosine bases—imprinted on the structure of the DNA. Here the DNA was acting as the substrate for a layer of information to be written on. This has nothing to do with the base sequence itself—the genetic code—it is defined as a level of epigenetic information impressed on the genetic level.
Epigenetics and Lamarckism
Even more-dramatic examples that violated classical Darwinism were soon uncovered.
|
Toward the end of World War II, a German-imposed food embargo in western Holland—a densely populated area already suffering from scarce food supplies, ruined agricultural lands, and the onset of an unusually harsh winter—led to the death by starvation of some 30,000 people. Detailed birth records collected during that so-called Dutch Hunger Winter have provided scientists with useful data for analyzing the long-term health effects of prenatal exposure to famine. Not only have researchers linked such exposure to a range of developmental and adult disorders, including low birth weight, diabetes, obesity, coronary heart disease, breast and other cancers, but at least one group has also associated exposure with the birth of smaller-than-normal grand-children. The finding is remarkable because it suggests that a pregnant mother's diet can affect her health in such a way that not only her children but her grandchildren (and possibly great-grandchildren, etc.) inherit the same health problems. In another study, unrelated to the Hunger Winter, researchers correlated grandparents' prepubertal access to food with diabetes and heart disease. In other words, you are what your grandmother ate. But, wait, wouldn't that imply what every good biologist knows is practically scientific heresy: the Lamarckian inheritance of acquired characteristics?[7] |
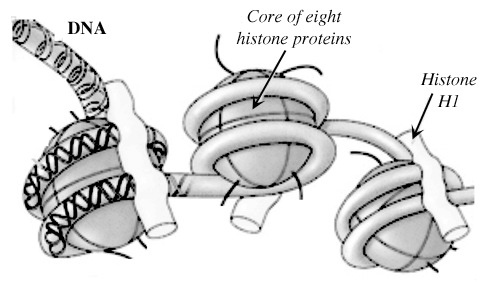
In this case, the epigenetic information involved chemical tagging of the histones, the protein 'spools' on which the foot-long DNA molecules are wrapped around to keep them manageable. This is an image of how histones and DNA combine:[8]
Reversible and site-specific histone modifications occur at multiple sites through acetylation of the histone proteins. It would seem—and this is currently an active area of research—that there is a connection between the epigenetic information written on the DNA and that written on the histones working in complementary directions: Methylation of DNA turns it off while acylation of histones turns them on. It seems that methylated DNA on non-acylated histones is hard to unwrap, so its information cannot be easily accessed, while un-methylated DNA on acylated histones is easy to unwrap and its information is more easily accessed. This diagram is a summary of what is currently known about the mechanism of epigenetic inheritance:[9]
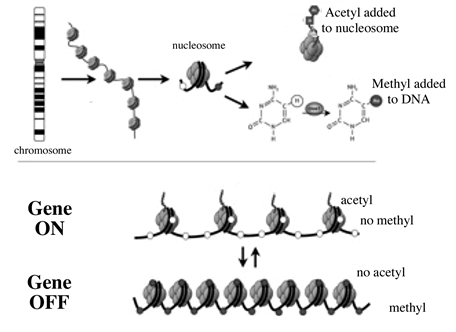
This mechanism of storing information about the current state of the organism is now well established; there are probably others at work as well.
Most of the investigations into epigenetic mechanisms are currently focused on medicine and the state of disease such as cancer and the like. There has not been much work on how this field impacts the mechanisms of evolution but it is clear that a new principle is involved: Information about the current state of the organism is imprinted on the genetic heritage and can be accumulated over the ages as it is passed on down a lineage.
|
The field of epigenetics has gained great momentum in recent years and is now a rapidly advancing field of biological and medical research. Epigenetic changes play a key role in normal development as well as in disease... The molecular mechanisms and biological proces¬ses in which epigenetic modifications play a primordial role [may] potential[ly] transfer epigenetic information across generations.[10] |
Recombination and Sex
Recombination, or "crossing over" as it is otherwise called, is occurs in the generation of the sex cells where two copies of a paternal chromosome and two copies of a maternal chromosome (i.e. 8 strands of DNA) entangle and crossover their genetic material.
|
Chromosomal crossover (or crossing over) is an exchange of genetic material between homologous chromosomes. It is one of final phases of genetic recombination, which occurs during prophase 1 of meiosis in a process called synapsis. Synapsis begins before the synaptonemal complex develops, and is not completed until near the end of prophase 1. Crossover usually occurs when matching regions on matching chromosomes break and then reconnect to the other chromosome.[11] |
It is this mixing of the genetic material that is at the heart of sexual reproduction and, as only sexual species evolve and spin off daughter species, can be considered to have a central in evolution.
In Darwinism, this process of breaking and reconnecting the DNA is considered random even though it is well established that there are 'hot-spots' (where crossing over occurs with a high frequency) and 'cold spots' (where crossing over never occurs).
Epigenetics and Recombination
Evidence is accumulating that there is a link between 'short term' epigenetic information and recombination with its long-term consequences. This is from the report of a biology convention in 2006:[12]
|
Carmen Sapienza (Temple University Medical School, Philadelphia, USA) reported that imprinted regions in humans are historical hotspots of recombination. Together with specific DNA sequences, epigenetic factors may have an important influence on the rate of meiotic recombination and the position of cross-overs. Using in silico and in vitro analyses, Sapienza's group have shown a relationship between increased rates of meiotic recombination and genomic imprinting. Imprinted regions showed more linkage disequilibrium, and had a significantly higher number of small haplotype blocks, than the non-imprinted regions. Their findings suggest that several factors, including both specific DNA sequences and epigenetics, are involved in controlling meiotic recombination in humans.[13] |
Other groups have also established connections between epigenetics and the rearrangement of the genome in recombination including the arabidopsis plant,[14] in humans[15], in the centromeres that control the structure of the eukaryote cell[16], and in the recombination that underlies the antibody diversity in the immune system.[17]
Why Sex?
One of the open questions in modern biology is: "Why sex?" The overwhelming preponderance of sexual reproduction in multicellular organisms is a puzzle because asexual reproduction is so much more efficient at generating progeny.
It is a well-known fact that, while Darwin titled his epochal work, "The Origin of Species," he did not actually propose any mechanism for the emergence of new species. The ideas he proposed, at best, dealt with the origin of races within a species, not new species themselves. To this day, there is no consensus as to how this happens, other than by a process of gradual divergence and gradual infertility between races.
This does accord with what is known, however, as illustrated by the human race. There has certainly been a lot of epigenetic learning and writing to genetic memory in the many tens of thousands of years since the first humans emerged in Africa. These are the human variants we call races.
|
The first theory, known as the "Out of Africa" model, is that Homo sapiens developed first in Africa and then spread around the world between 100 and 200,000 years ago, superseding all other hominid species. The implication of this argument is that all modern people are ultimately of African descent. The other theory, known as the "Multi-regional" Model, is that Homo sapiens evolved simultaneously in different parts of the world... Although the debate is far from concluded, it is probably fair to say that the bulk of scientists support the Out of Africa hypothesis and believe that all humans share a common origin.[18] |
Examples of the innovations expressed in the emergence of the human races are the ability to digest milk through adulthood (a rarity in the stay-at-home Africans; common in Europeans) and the loss of UV-protecting but vitamin D discouraging melanin in the races in sun-deprived northern latitudes. For all these epigenetic and genetic changes, however, the ability of Black Africans and White Europeans to interbreed is in no way diminished.[19] In fact, a quite-opposite phenomenon is firmly established in biology: that of hybrid vigor: "An increase in the performance of hybrids over that of purebreds, most noticeably in traits such as fertility and survivability."[20]
In the theory presented here, the epigenetic-directed recombination of genetic material is the key mechanism of speciation and resultant reproductive isolation. Evidence that this might be correct is to be found in the rather odd sequence of events leading up to the formation of the haploid sex cells (with one set of chromosomes) from the diploid germ cells (with two sets of chromosomes). One obvious reason for this haploid-diploid alternation is to prevent a buildup of chromosome number that would happen if the sex cells were diploid: the children would have four sets, the grandchildren eight, the great-grandchildren sixteen, etc.
The obvious way to get two haploid cells from a diploid cell would be to have a regular cell division (mitosis) that skips the chromosome duplication step. This is not the case. The formation of the sex cells (meiosis) adds a seemingly unnecessary step that just adds to the workload. First the two sets of chromosomes—the paternal set and the maternal set—are duplicated. The cell now has four sets of chromosomes! These all commingle into what is called the tetraplex or synaptic complex,[21] the stage when recombination and reorganization of the genetic material occurs. The four sets of chromosomes are now progressively reduced to one set by two rounds of cell division to create four haploid sex cells.
Current biology has no good rationale for this complicated way of doing things, as recombination is considered to be random chance-and-accident. In the perspective developed here, however, this abundance of chromosomes hints at some currently-uncharacterized mechanism for the directed reorgani¬zation of genetic material while also ensuing that the new daughter species can be 'brought to term' successfully by the mother species.[22]
If epigenetic-directed recombination turns out to be at the heart of speciation, it would provide a simple answer as to why almost all species are sexual: only sexual species can evolve; only sexual species can give rise to new species and more sophisticated organisms. The adoption of the asexual mode of reproduction, while advantageous in the moment, is an evolutionary dead end.
|
The evolution of traditional, female-only asexuality typically leads to a swift extinction. We know this because although such species frequently evolve, they don't stay around for long. If you look at the tree of life, female-only asexual groups are all out on the twigs: there are no great asexual lineages equivalent to fish or birds. Instead, the asexual groups are a few species of snail here, a dandelion there.[23] |
Without sex, the highest forms of life would be the simple unicellular forms that predominated the first billion years of life on earth. There would have been no Cambrian Explosion of multi-cellular forms, and certainly no humans.
Unification Thought puts sexuality at the very center of human life (and the Fall); if this perspective has any validity it would seem that Molecular Biology should have sex as the dynamo of evolution.
Logos, Learning and Evolution
We have now discussed this sequence of cause and effect:
- The analog form and function of the organism resonates with the Logos (sophisticated natural law) and learns about new aspects of the structure of the Logos.
- These insights into the structure of the Logos are stored as epigenetic information written on the DNA inherited from the ancestors and the combination passed on to the descendants.
- This incremental information guides small changes in the recombination events in the formation of the sexual gametes of the species. This is microevolution guided by input from the Logos.
- The accumulation of epigenetic information in the lineage reaches a saturation point and it is used to guide a massive recombinatorial reorgan¬ization of the genetic material. This is macroevolution guided by input from the Logos, and it results in a new daughter species whose genetic material is incompatible with that of the mother species. As all the short-term epigenetic information has now been incorporated into the long-term genetic material, it is discarded and the result is a "spotless" DNA with a fresh new start—a blank page to start anew accumulating wisdom from the Logos.
- Evolution is the incremental accumulation of wisdom about the Logos being passed on down a lineage: first in short-term epigenetic memory, then in long-term genetic memory.
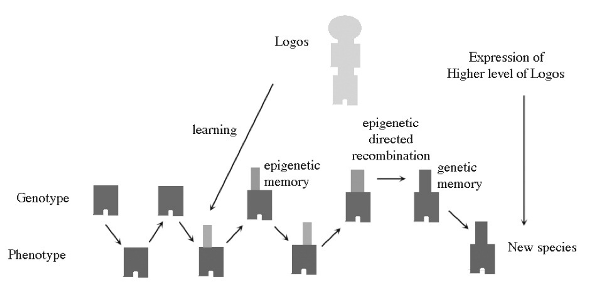
An example of radical rearrangement of the genome is to be found in the lineage leading to Adam and Eve. The human genome has one less chromosome than all of our primate cousins. It is now well-established that the second-largest human chromosome is a result of the fusion of two smaller great ape chromosomes:
|
All great apes apart from man have 24 pairs of chromosomes. There is therefore a hypothesis that the common ancestor of all great apes had 24 pairs of chromosomes and that the fusion of two of the ancestor's chromosomes created chromosome 2 in humans.... Evidence for fusing of two ancestral chromosomes to create human chromosome 2 and where there has been no fusion in other Great Apes is: (1) The analogous chromosomes (2p and 2q) in the non-human great apes can be shown, when laid end to end, to create an identical banding structure to the human chromosome 2. (2) The remains of the sequence that the chromosome has on its ends (the telomere) is found in the middle of human chromosome 2 where the ancestral chromosomes fused. (3) The detail of this region... is exactly what we would expect from a fusion. (4) This telomeric region is exactly where one would expect to find it if a fusion had occurred in the middle of human chromosome 2. (5) The centromere of human chromosome 2 lines up with the chimp chromosome 2p chromosomal centromere. (6) At the place where we would expect it on the human chromosome, we find the remnants of the chimp 2q centromere. Not only is this strong evidence for a fusion event, but it is also strong evidence for common ancestry; in fact, it is hard to explain by any other mechanism.[24] |
Darwinists would claim that this fusion of chromosomes was a fortuitous and accidental event during recombination; the Unification view is that this was an event guided by input from the Logos with the goal of creating the human form.
Recording the Wave
Current evolutionary science is constructed on the foundation of classical physics which considered only the observable external aspects of matter in its theories. Modern physics, however, has been forced by experiment to conclude that the external aspects are insufficient, and that an internal, unobservable aspect has to be taken into account to explain things. This is the wave-function of quantum physics (also called 'probability amplitude' among other names).
The concepts 'external' and 'internal' have a precise mathematical definition in the new physics: The external aspects are described with real numbers, while the internal aspects are described with complex numbers. Real numbers have a single, linear component, while complex numbers have two components, one linear and one rotational. It is the relationship between these two types of numbers that mathematically describes the relationship between the sung sang and hyung sang aspects in the Divine Principle.
In classical science, the focus is on particles and waves play only a supporting role. In a modern, unified science, the properties of waves play a central role. Understanding of waves is most highly developed in the discipline of music, and it is there that we can isolate the concept of a 'wave-in-full' and its relation to a 'wave-in-image.'
Wave in Full and Wave in Image
Consider an orchestra and chorus performing the soaring climax to Beethoven's Ode to Joy in Carnegie Hall. The singers and musicians are each reading their section of the digital score and belting out the corresponding analog sound wave on their instruments. All of these sound-wave generators fill the hall's million-or-so cubic feet with an extraordinarily complex, slowly changing (on the timescale of sound) standing wave of air pressure that structures the air in the hall. We shall refer to it as the analog 3-D 'wave-in-full' of the Ode to Joy.
A microphone locally samples the changing air pressure and creates a 1-D analog wave of electrons in a wire that is amplified and recorded on a tape. This analog wave is a 'wave-in-image' of the Ode to Joy. It may be further converted into digital form for a CD recording. Playing the tape containing that recorded wave-in-image or playing a CD containing the digital form simply reverses the process to make a new wave-in-full.
Our experience of Beethoven's symphony is not actually that of the wave-in-full. Our eardrum converts it into an analog wave-in-image, which the cochlea in turn converts into digital form for transmission along the auditory nerve to the brain, where the wave-in-full in recreated in our minds. Our two ears pick up slightly different wave-in-images which allows the brain to recreate more accurately the wave-in-full in our minds.
To summarize, these are the basic concepts:
- A set of musical instruments generate waves that structure the air in a concert hall (wave-in-full)
- A linear readout of this is captured for recording (wave-in-image)
- The wave is sampled by an electronic device and its result recorded as a series of numbers (analog to digital)
- The process is reversed in playback (digital to analog, wave-in-image to wave-in-full)
These concepts about waves can be fruitfully applied to living systems, which can be considered as structured water:
- A set of proteins generate waves that structures the water in a cell (wave-in-full)
- A linear readout of this is captured for recording (wave-in-image)
- The wave is sampled by proto t-RNA and its result recorded as a series of numbers (analog to digital)
- The process is reversed in playback (digital to analog, wave-in-image to wave-in-full.
This brief summary of the wave aspect is treated more fully in another paper by this author.[25]
Memory
With the concepts of wave-in-full and wave-in-image, we can speculate that there are other mechanisms involved in epigenetics other than just the control of recombination, mechanisms that result in new information being inserted into the genome. One possibility is a primordial mechanism that predates the origin of the triplet code method of protein synthesis. In this view, the shape of the pre-transfer RNAs (p-tRNA) acts as the transducer between the analog realm of waves and form, and the digital information realm of RNA and DNA. This is illustrated in the diagram:
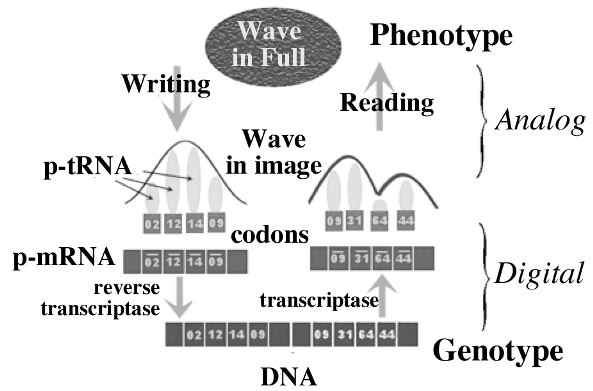
Supporting evidence for this suggestion are:
1. Forms of life based on RNA predated the RNA-DNA-protein world. The process illustrated above would have predated the triplet code, and would have served as the precursor for its emergence.
|
For decades, many researchers thought that ribonucleic acid, or RNA, was nothing more than a molecular interpreter that helps translate DNA codes into proteins. But research over the past 15 years... has been lending credence to the notion of a so-called "RNA world," an era in early evolution when all life forms were based on RNA.[26] |
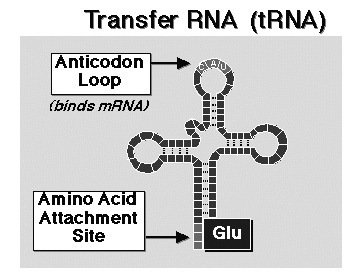 2. The set of 64 codon-bearing transfer RNA have a variety of shapes that have little or no connection to the amino-acids they carry in their role in the triplet code synthesis of pro¬teins. They, or a simpler precursor, are thus well-suited for their putative role as adaptors be-tween the analog and digital realms. This is a diagram of the folded shape of one tRNA molecule:[27]
2. The set of 64 codon-bearing transfer RNA have a variety of shapes that have little or no connection to the amino-acids they carry in their role in the triplet code synthesis of pro¬teins. They, or a simpler precursor, are thus well-suited for their putative role as adaptors be-tween the analog and digital realms. This is a diagram of the folded shape of one tRNA molecule:[27]
3. The human genome has over 300,000 genes that are similar to reverse transcriptase. Current thinking is that they are useless remnants of ancestral viral invasions similar to retroviral HIV. They are not thought to have a constructive role.
|
Over 11% of the mammalian genome is composed of retrovirus-like [DNA sequences]: 'transposable elements in which transposition involves a process of reverse transcription with an RNA intermediate similar to that of a retrovirus'. Compare this with only ~2.5% of the human genome which encodes unique (non-repeated) genes! ...Another type of repetitive DNA element consists of retrovirus-like elements, or human endogenous retroviruses, representing about 7% of the human genome. Their structure closely resembles that of retroviruses, carrying internal sequences with homology to [reverse transcriptases] flanked by long terminal repeats. Similar sequences occur in all organisms, from yeast to vertebrates.[28] |
The thesis presented here suggests that these elements play an important, if only occasionally invoked, role in cell memory and the long-term development of the genome.
4. The ability of viral RNA (or DNA which is first copied into RNA) to suborn the workings of a cell is well-established. It is not, therefore, too much of a speculation to think that endogenous RNA recalled from memory can also direct the form and function of the cell. In this view, the viral RNA takes over the fundamental role of memory RNA (not directly connected to a role in protein synthesis), not as is currently thought, by a fortuitous combination of circumstances. While DNA (and its random modifications) plays a central role in Darwinism and the modern synthesis; it is more and more being assigned a passive role where its information content is under the control of RNA:
|
Many RNAs are involved in modifying other RNAs. Introns are spliced out of pre-mRNA by spliceosomes, which contain several small nuclear RNAs (snRNA), or the introns can be ribozymes that are spliced by themselves. RNA can also be altered by having its nucleotides modified to other nucleotides than A, C, G and U. In eukaryotes, modifications of RNA nucleotides are generally directed by small nucleolar RNAs (snoRNA), found in the nucleolus and cajal bodies. snoRNAs associate with enzymes and guide them to a spot on an RNA by base pairing to that RNA.[29] |
This discussion does suggest that the nucleolus—a dense congregation of a multitude RNA molecules in the nucleus—plays a much more central role in the running of the cell than the simple 'housekeeping' role it is now assigned: that of making ribosomes for export to the cytoplasm where they construct the proteins.
Conclusion
Darwin's insight into the evolution of life from simple to sophisticated can be summed up as "variation in a lineage followed by natural selection." The development of this into the current amalgam of genetics and molecular biology called the Modern Synthesis embraced the atheistic concept that the variation was totally random; there was no inherent design that was being expressed over time.
The view presented here, while embracing much of what is firmly established in the Modern Synthesis, denies the random no-design aspect that has been imposed on science by atheistic thinkers.
Variation is not random; it involves learning about the Logos and a deposition of this wisdom in the mainly-digital information that is accumulated and passed on down the lineage. It can be considered an ultra-modern synthesis of Molecular biology, Darwinism and Lamarckism. It also echoes some of the core principles of Unificationism without any imposition of philosophical bias.
The current understanding of evolution is based on the Central Dogma: that the flow of information is one-way only, from genotype to phenotype. We have shown that there is good reason to suspect that this dogma is false, that information is passed from the phenotype to the genotype, initially as short-term epigenetic storage, then as long-term genetic storage.
Notes
[1] Sun Myung Moon, Cheon Seong Gyeong, Book 4, Chapter 10, Section 2: Adam's Family Fell into Satan's Possession. At: http://www.tparents.org/Moon-Books/SunMyungMoon-CSG/CSG-04-10-02.htm
[2] http://www.pbs.org/wgbh/nova/sciencenow/3411/02.html
[3] http://www.sciencedaily.com/releases/2009/05/090518111723.htm
[4] A. Otani, Beyond Darwinism: Towards Unification Science (Tokyo: UTI, 2009).
[5] http://www.eskimo.com/~billb/weird/end.html
[6] http://www.nature.com/scitable/topicpage/Genomic-Imprinting-and-Patterns-of-Disease-Inheritance-899
[7] https://notes.utk.edu/Bio/greenberg.nsf/0/b360905554fdb7d985256ec5006a7755
[8] http://www.newgeology.us/HistoneH1.jpg
[9] http://cnx.org/content/m26565/latest/
[10] Review on Google Books of Jörg Tost, Epigenetics (Norfolk, UK: Caster Academic Press, 2008).
[11] http://en.wikipedia.org/wiki/Chromosomal_crossover
[12]Advances in the Genetics and Epigenetics of Gene Regulation and Human Disease: A Report on the Human Genome Organisation (HUGO), 11th Human Genome Meeting, Helsinki, Finland, 31 May-3 June 2006.
[13] Http://genomebiology.com/2006/7/8/325
[14] Haibo Yin, Xia Zhang, Jun Liu, Youqun Wang, Junna He, Tao Yang, Xuhui Hong, Qing Yang, and Zhizhong Gong, "Epigenetic Regulation, Somatic Homologous Recombination and Abscisic Acid Signaling Are Influenced by DNA Polymerase Epsilon Mutation in Arabidopsis," Plant Cell 21/2 (February 2009): 386-402.
[15] Martin I. Sigurdsson, Albert V. Smith, Hans T. Bjornsson and Jon J. Jonsson, "HapMap Methylation-Associated SNPs, Markers of Germline DNA Methylation, Positively Correlate with Regional Levels of Human Meiotic Recombination," Genome Research 19/4 (26 April 2009): 581-589.
[16] http://www.epidna.com/showabstract.php?pmid=18541703
[17] http://www.reeis.usda.gov/web/crisprojectpages/207316.html
[18] http://www.bbc.co.uk/worldservice/specials/1624_story_of_africa/page92.shtml
[19] Experimentation in this area has begun. For example, see "Hybrid Vigor and Transgenerational Epigenetic Effects on Early Mouse Embryo Phenotype." http://www.biolreprod.org/content/79/4/638.abstract
[20] http://www.alpacas.com/AlpacaLibrary/GlossaryGL.aspx
[21] http://www.nature.com/nrm/journal/v4/n11/glossary/nrm1241_glossary.html
[22] I have speculated on a possible mechanism for such a speciation mechanism in my book "Do Proteins Teleport in an RNA World." A gratis copy can be obtained from the UTI office or by sending an email request with your postal address to the author at: This email address is being protected from spambots. You need JavaScript enabled to view it..
[23] http://judson.blogs.nytimes.com/2008/09/23/evolving-the-single-daddy/
[24] Alec MacAndrew, Human Chromosome 2 is a Fusion of Two Ancestral Chromosomes. http://www.evolutionpages.com/chromosome_2.htm
[25] Richard Lewis, "Dual Characteristics in Unification Thought and Quantum Science." A copy of this paper in the PDF format can be obtained from the author by sending a request to: This email address is being protected from spambots. You need JavaScript enabled to view it.
[26] http://www.sciencedaily.com/releases/1998/09/980917081316.htm
[27] http://universe-review.ca/F11-monocell.htm
[28] http://www.microbiologybytes.com/virology/Retroviruses.html
[29] http://en.wikipedia.org/wiki/RNA#In_RNA_processing